
User:Marcus chatfield/sandbox
 | This is a user sandbox of Marcus chatfield. You can use it for testing or practicing edits. This is not the sandbox where you should draft your assigned article for a dashboard.wikiedu.org course. To find the right sandbox for your assignment, visit your Dashboard course page and follow the Sandbox Draft link for your assigned article in the My Articles section. |
Overview
[edit]Historically the ocean and terrestrial forest ecosystems have been the major natural Carbon (C) sinks. New research on the role of vegetated coastal ecosystems have highlighted their potential as highly efficient C sinks, and led to the scientific recognition of the term “Blue Carbon"[1]. Although the ocean’s vegetated habitats cover less than 0.5% of the seabed, they are responsible for more than 50%, and potentially up to 70% of all carbon storage in ocean sediments[1]. Mangroves, salt marshes and seagrasses make up the majority of the ocean’s vegetated habitats, but only equal 0.05% of the plant biomass on land. Despite their small footprint, they can store a comparable amount of carbon per year and are highly efficient carbon sinks. Seagrasses, mangroves and salt marshes can capture CO2 from the atmosphere by sequestering the C in their underlying sediments, in underground and belowground biomass, and in dead biomass[2]. In plant biomass like leaves, stems, branches or roots, blue carbon can be sequestered for years to decades, and for thousands to millions of years in underlying plant sediments. Current estimates of long-term blue carbon C burial capacity are variable, and research is ongoing[2]. Although vegetated coastal ecosystems cover less area and have less aboveground biomass than terrestrial plants, they have the potential to impact longterm C sequestration, particularly in sediment sinks[1]. One of the main concerns with Blue Carbon is the rate of loss of these important marine ecosystems is much higher than any other ecosystem on the planet, even compared to rainforests. Current estimates suggest a loss of 2-7% per year, which is not only lost carbon sequestration, but also lost habitat that is important for managing climate, coastal protection, and health[1].
Types of blue carbon ecosystems
[edit]Seagrass
[edit]Seagrass are a group of about 60 angiosperm species that have adapted to an aquatic life, and can grow in meadows along the shores of all continents except Antarctica[3]. Seagrass meadows form in maximum depths of up to 50m, depending on water quality and light availability, and can include up to 12 different species in one meadow[3]. These seagrass meadows are highly productive habitats that provide many ecosystem services, including sediment stabilization, habitat and biodiversity, better water quality, and carbon and nutrient sequestration[4]. The current documented seagrass area is 177,000 km2, but is thought to underestimate the total area since many areas with large seagrass meadows have not been thoroughly documented[3]. Most common estimates are 300,000 to 600,000 km2, with up to 4,320,000 km2 suitable seagrass habitat worldwide[5]. Although seagrass makes up only 0.1% of the area of the ocean floor, it accounts for approximately 10-18% of the total oceanic carbon burial[6]. Currently global seagrass meadows are estimated to store as much as 19.9 Pg (gigaton, or billion tons) of organic carbon[6].Carbon primarily accumulates in marine sediments, which are anoxic and thus continually preserve organic carbon from decadal-millennial time scales. High accumulation rates, low oxygen, low sediment conductivity and slower microbial decomposition rates all encourage carbon burial and carbon accumulation in these coastal sediments[3]. Compared to terrestrial habitats that lose carbon stocks as CO2 during decomposition or by disturbances like fires or deforestation, marine carbon sinks can retain C for much longer time periods. C sequestration rates in seagrass meadows vary depending on the species, characteristics of the sediment, and depth of the habitats, but on average the carbon burial rate is approximately 138 g C m-2 yr-1[2]. Seagrass habitats are threatened by coastal eutrophication, increased sedimentation and coastal development[2], as well as sea-level rise which may decrease light availability for photosynthesis, and increased seawater temperatures[3]. Seagrass loss have accelerated over the past few decades, from 0.9% per year prior to 1940 to 7% per year in 1990, with about 1/3 of global loss since WWII[7]. Scientists encourage protection and continued research of these ecosystems for organic carbon storage, valuable habitat and other ecosystem services.
Mangrove
[edit]Mangroves are woody halophytes that form intertidal forests and provide many important ecosystem services including coastal protection, nursery grounds for coastal fish and crustaceans, forest products, recreation, nutrient filtration and carbon sequestration[8].Currently they are found in 123 countries, with 73 identified species[9]. They grow along coastlines in subtropical and tropical waters, depending mainly on temperature, but also vary with precipitation, tides, waves and water flow[10]. Because they grow at the intersection between land and sea, they have semi-terrestrial and marine components, including unique adaptations including aerial roots, viviparous embryos, and highly efficient nutrient retention mechanisms[11]. Mangroves cover approximately 150,000 km2 worldwide, but have declined by 20% in the last 25 years, mainly due to coastal development and land conversion. Mangrove deforestation is slowing, from 1.04% loss per year in the 1980s to 0.66% loss in the early 2000s[9], as research and understanding of mangrove benefits have increased. Mangrove forests are responsible for approximately 10% of global carbon burial[12], with an estimated carbon burial rate of 174 g C m-2 yr-1[11]. Like seagrass, mangroves have potential for high levels of carbon sequestration, and account for 3% of the global carbon sequestration by tropical forests, and 14% of the global coastal ocean's carbon burial[10]. Mangroves are naturally disturbed by floods, tsunamis, coastal storms like cyclones and hurricanes, lightning, disease and pests, and changes in water quality or temperature[11]. Although they are resilient to many of these natural disturbances, they are highly susceptible to human impacts, including urban development, aquaculture, mining, and overexploitation of shellfish, crustaceans, fish and timber[9][11]. This is an important habitat to conserve and repair when possible, for globally important ecosystem services and carbon sequestration.
Marsh
[edit]Marshes, or tidal wetlands are intertidal ecosystems dominated by herbaceous vegetation, found globally on coastlines from arctic to subtropical waters, where they are replaced by mangroves as the dominant coastal vegetation[13]. Marshes have high productivity, with a large portion of primary production in belowground biomass, which can form deposits up to 8m deep[13]. Marshes provide valuable habitat for plants, birds, and other animals, safe refuges for juvenile fish, protect coastal habitat from storm surge and flooding, and can reduce nutrient loading to coastal waters[14]. Additionally, organic sedimentation allows marshes to serve as carbon sinks, since they are dominated by anaerobic decomposition that promotes C sequestration in underground biomass, similarly to mangrove and seagrass habitats[15]. Salt marshes cover approximately 22,000 to 400,000 km2 globally, with an estimated carbon burial rate of 210 g C m-2 yr-1[13]. Tidal marshes have been impacted by humans for centuries, including modification for grazing, haymaking, reclamation for agriculture, development and ports, evaporation ponds for salt production, modification for aquaculture, insect control, tidal power and flood protection[16]. Marshes are also susceptible to pollution from oil, industrial chemicals, and most commonly, eutrophication. Introduced species, sea-level rise, river damming and decreased sedimentation are additional longterm changes that affect marsh habitat, and in turn, may affect carbon sequestration potential[16].
Algae
Both macroalgae and microalgae are being investigated as possible means of carbon sequestration[17].[18][19][20] Because algae lack the complex lignin associated with terrestrial plants, the carbon in algae is released into the atmosphere more rapidly than carbon captured on land[21].[19] Algae have been proposed as a short-term storage pool of carbon that can be used as a feedstock for the production of various biogenic fuels. Microalgae are often put forth as a potential feedstock for carbon-neutral biodiesel and biomethane production due to their high lipid content.[17] Macroalgae, on the other hand, do not have high lipid content and have limited potential as biodiesel feedstock, although they can still be used as feedstock for other biofuel generation.[19] Macroalgae have also been investigated as a feedstock for the production of biochar. The biochar produced from macroalgae is higher in agriculturally important nutrients than biochar produced from terrestrial sources.[20] Another novel approach to carbon capture which utilizes algae is the Bicarbonate-based Integrated Carbon Capture and Algae Production Systems (BICCAPS) developed by a collaboration between Washington State University in the United States and Dalian Ocean University in China. Many cyanobacteria, microalgae, and macroalgae species can utilize carbonate as a carbon source for photosynthesis. In the BICCAPS, alkaliphilic microalgae utilize carbon captured from flue gases in the form of bicarbonate[22][23]. In South Korea, macroalgae have been utilized as part of a climate change mitigation program. The country has established the Coastal CO2 Removal Belt (CCRB) which is comprised of artificial and natural ecosystems. The goal is to capture carbon using large areas of kelp forest.[24]
Ecosystem restoration
Restoration of mangrove forests, seagrass meadows, marshes, and kelp forests has been implemented in many countries[25][26][27]. These restored ecosystems have the potential to act as carbon sinks. Restored seagrass meadows were found to start sequestering carbon in sediment within about four years. This was the time needed for the meadow to reach sufficient shoot density to cause sediment deposition.[27] Similarly, mangrove plantations in China showed higher sedimentation rates than barren land and lower sedimentation rates than established mangrove forests. This pattern in sedimentation rate is thought to be a function of the plantation’s young age and lower vegetation density.[26]
Restoration of mangrove forests, seagrass meadows, marshes, and kelp forests has been implemented in many countries[28][29][30]. These restored ecosystems have the potential to act as carbon sinks. Restored seagrass meadows were found to start sequestering carbon in sediment within about four years. This was the time needed for the meadow to reach sufficient shoot density to cause sediment deposition.[30] Similarly, mangrove plantations in China showed higher sedimentation rates than barren land and lower sedimentation rates than established mangrove forests. This pattern in sedimentation rate is thought to be a function of the plantation’s young age and lower vegetation density.[29]
Nutrient stoichiometry of seagrasses
[edit]The primary nutrients determining sea grass growth are carbon, nitrogen, phosphorous, and light for photosynthesis. N and P can be acquired from sediment pore water or from the water column, and sea grasses can uptake both N in both ammonium (NH4+) and nitrate (NO3-) form.[13]
A number of studies from around the world have found that there is a wide range in the concentrations of C, N, and P in seagrasses depending on their species and environmental factors. For instance, plants collected from high-nutrient environments had lower C:N and C:P ratios than plants collected from low-nutrient environments. Sea grass stoichiometry does not follow the Redfield ratio commonly used as an indicator of nutrient availability for phytoplankton growth. In fact, a number of studies from around the world have found that the proportion of C:N:P in sea grasses can vary significantly depending on their species, nutrient availability, or other environmental factors. Depending on environmental conditions, sea grasses can be either P-limited or N-limited. [31]
An early study of sea grass stoichiometry suggested that the “Redfield” balanced ratio between N and P for sea grasses is approximately 30:1.[32] However, N and P concentrations are strictly not correlated, suggesting that sea grasses can adapt their nutrient uptake based on what is available in the environment. For example, sea grasses from meadows fertilized with bird excrement have shown a higher proportion of phosphate than unfertilized meadows. Alternately, sea grasses in environments with higher loading rates and organic matter diagenesis supply more P, leading to N-limitation. P availability in T. testudinum is the limiting nutrient. The nutrient distribution in T. testudinum ranges from 29.4-43.3% C, 0.88-3.96% N, and 0.048-0.243% P. This equates to a mean ratio of 24.6 C:N, 937.4 C:P, and 40.2 N:P. This information can also be used to characterize the nutrient availability of a bay or other water body (which is difficult to measure directly) by sampling the sea grasses living there.[16]
Light availability is another factor that can affect the nutrient stoichiometry of sea grasses. Nutrient limitation can only occur when photosynthetic energy causes grasses to grow faster than the influx of new nutrients. For example, low light environments tend to have a lower C:N ratio.[16] Alternately, high-N environments can have an indirect negative effect to sea grass growth by promoting growth of algae that reduce the total amount of available light.[13] </ref>
Nutrient variability in sea grasses can have potential implications for wastewater management in coastal environments. High amounts of anthropogenic nitrogen discharge could cause eutrophication in previously N-limited environments, leading to hypoxic conditions in the sea grass meadow and affecting the carrying capacity of that ecosystem.[16]</ref>
A study of annual deposition of C, N, and P from P. Oceanica sea grass meadows in northeast Spain found that the meadow sequestered 198 g C m-2 yr-1, 13.4 g N m-2 yr-1, and 2.01 g P m-2 yr-1 into the sediment. Subsequent remineralization of carbon from the sediments due to respiration returned approximately 8% of the sequestered carbon, or 15.6 g C m-2 yr -1.[33]
Distribution and decline of blue carbon ecosystems
[edit]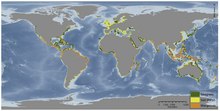
Seagrasses, mangroves and marshes are types of vegetated coastal habitats that cover approximately 49 million hectares worldwide [35].Seagrass ecosystems range from polar to tropical regions, mangroves are found in tropical and sub-tropical ecosystems and tidal marshes are found in mostly temperate regions such as on the east coast of the United States [36]. As habitats that sequester carbon are altered and decreased, that stored amount of C is being released into the atmosphere, continuing the current accelerated rate of climate change. Impacts on these habitats globally will directly and indirectly release the previously stored carbon, which had been sequestered in sediments of these habitats. Declines of vegetated coastal habitats are seen worldwide; examples seen in mangroves are due to clearing for shrimp ponds such is the case in Indonesia, while in seagrasses there are both natural causes due to pathogens and may be exacerbated by anthropogenic effects. Quantifying rates of decrease are difficult to calculate, however measurements have been estimated by researchers indicating that if blue carbon ecosystems continue to decline, for any number of reasons, 30-40% of tidal marshes and seagrasses and approximately 100% of mangroves could be gone in the next century[37].
Decline in seagrasses are due to a number of factors including drought, water quality issues, agricultural practices, invasive species, pathogens, fishing and climate change [38].Over 35% of global mangrove habitat remains. Decreases in habitat is due to damming of rivers, clearing for aquaculture, development etc., overfishing, and climate change, according to the World Wildlife Fund[39]. Nearly 16% of mangroves assessed by the IUCN are on the IUCN Red List; due to development and other causes 1 in 6 worldwide mangroves are in threat of extinction [40]. Dams threaten habitats by slowing the amount of freshwater reaching mangroves. Coral reef destruction also plays a role in mangrove habitat health as reefs slow wave energy to a level that mangroves are more tolerant of. Salt marshes may not be expansive worldwide in relation to forests, but they have a C burial rate that is over 50 times quicker than tropical rainforests. Rates of burial have been estimated at up to 87.2 ± 9.6 Tg C yr-1 which is greater than that of tropical rainforests,53 ± 9.6 Tg C yr-1 [41]. Since the 1800s salt marshes have been disturbed due to development and a lack of understanding their importance. The 25% decline since that time has led to a decrease in potential C sink area coupled with the release of once buried C. Attributed to degraded marsh habitat will be a decrease in C stock in sediments, a decrease in plant biomass and thus a decrease in photosynthesis reducing the amount of CO2 taken up by the plants, C in plant blades not transferring into the sediment, and lastly erosive processes could be accelerated due to the lack of plant biomass, accelerating buried C release to the atmosphere [42].
Reasons for decline of mangroves, seagrass, and marshes include land use changes, climate and drought related effects, dams built in the watershed, convergence to aquaculture and agriculture, land development and sea-level rise due to climate change. Increases in these activities can lead to significant decreases in habitat available and thus increases in released C from sediments. As anthropogenic effects and climate change are heightened, the effectiveness of blue carbon sinks will diminish and CO2 emissions will be further increased. Data on the rates at which CO2 is being released into the atmosphere is not robust currently, however research is being conducted to gather a better information to analyze trends. Loss of underground biomass (roots and rhizomes) will allow for CO2 to be emitted changing these habitats into sources rather than carbon sinks [43].
Sedimentation and blue carbon burial
[edit]Organic carbon is only sequestered from the oceanic system if it reaches the sea floor and gets covered by a layer of sediment. Reduced oxygen levels in buried environments mean that tiny bacteria who eat organic matter and respire CO2 can’t decompose the carbon, so it is removed from the system permanently. Organic matter that sinks, but is not buried by a sufficiently deep layer of sediment is subject to re-suspension by changing ocean currents, bioturbation by organisms that live in the top layer of marine sediments, and decomposition by heterotrophic bacteria. If any of these things occur, then the organic carbon is put back in the system. Carbon sequestration takes place only if burial rates by sediment are greater than the long term rates of erosion, bioturbation, and decomposition.[13]</ref>
Spatial variability in sedimentation
[edit]Sedimentation is the rate at which floating or suspended particulate matter sinks and accumulates on the ocean floor. The faster (more energetic) the current, the more sediment it can pick up. As sediment laden currents slow, the particles fall out of suspension and come to rest on the sea floor. In other words, a fast current can pick up lots of heavy grains, where as a slow current can pick up only tiny pieces. As one can imagine, different places in the ocean vary drastically when it comes to the amount of suspended sediment and rate of deposition.[13]
Open ocean
[edit]The open ocean has very low sedimentation rates because most of the terrigenous sediments that make it here are carried by the wind. Aeolian transport accounts for only a small fraction of the total sediment delivery to the oceans. Additionally, there is much less plant and animal life to be buried. Therefore, carbon burial rates are relatively slow in the open ocean.[16]
Coastal margins
[edit]Coastal margins have high sedimentation rates because of rivers, which account for the vast majority of sediment delivery to the ocean. In most cases, sediments are deposited near the river mouth or are transported in the alongshore direction due to wave forcing. In some places sediment falls into submarine canyons and is transported off-shelf, if the canyon is sufficiently large or the shelf is narrow. Coastal margins also contain diverse and plentiful marine species, especially in paces that experience periodic upwelling. More marine life plus higher sedimentation rates create hotspots for carbon burial.[44]
Submarine Canyons
[edit]Marine canyons are magnets for sediment, because as currents carry sediment on the shelf in the alongshore direction, the path of the current crosses canyons perpendicularly. When the same amount of water flow is suddenly in much deeper water it slows down, and deposits sediment. Due to the extreme depositional environment, carbon burial rates in the Nazare canyon near Portugal are 30x greater than the adjacent continental slope! This canyon alone accounts for about 0.03% of global terrestrial OC burial in marine sediments. This may not seem like much, but the Nazarre submarine canyon only makes up 0.0001% of the area of the worlds oceans.[16]</ref>
Human changes to global sedimentary systems
[edit]Humans have been modifying sediment cycles on a massive scale for thousands of years through a number of mechanisms.
Agriculture/land clearing
[edit]When humans started clearing land to grow crops, they ruined the ability of the land to retain sediment. In a natural ecosystem, roots from plants hold sediment in place when it rains. Trees and shrubs reduce the amount of rainfall that impacts the dirt, and create obstacles that forest streams must flow around. When all vegetation is removed rainfall impacts directly on the dirt, there are no roots to hold on to the sediment, and there is nothing to stop the stream from scouring banks as it flows straight downhill. For many thousands of years, the net effect of humans on global sediment cycles was an increase in sedimentation due to this process.
Dams
[edit]The first dams date back to 3000 BC and were built to control flood waters for agriculture. When sediment laden river flow reaches a dam’s reservoir, the water slows down as it pools. Since slower water can’t carry as much sediment, virtually all of the sediment falls out of suspension before the water passes through the dam. The result is that most dams are nearly 100% efficient sediment traps. For thousands of years, there were too few dams to have a significant impact on global sedimentary cycles. The popularization of hydroelectric power in the last century has caused an enormous boom in dam building. Currently only a third of the world’s largest rivers flow unimpeded to the ocean.[45]
Channelization
[edit]In a natural system, the banks of a river will meander back and forth as different channels erode, accrete, open, or close. Seasonal floods regularly overwhelm riverbanks and deposit nutrients on adjascent flood plains. These services are essential to natural ecosystems, but are quite troublesome for people. Humans love to build houses and developments close to rivers, but hate watching their buildings get flooded or tumble down a bank. In response rivers in populated areas are often channelized, meaning that their banks and sometimes beds are armored with a hard material that prevents erosion and fixes them in place. This prevents erosion because there is no soft substrate left for the river to take downstream.
Impacts
[edit]Currently, the net effect of humans on global sedimentary cycling is a drastic reduction in the amount of sediment that makes it to the ocean. If we continue to build dams and channelize rivers, we will start to see a number of problems in coastal areas including sinking deltas, shrinking beaches, and disappearing salt marshes. In addition, it’s possible that we might ruin one of the biggest carbon sinks that we know of. Without sequestration of carbon in coastal marine sediments, we will likely see accelerated global climate change.[46]
Factors influencing blue carbon burial rates
Density of vegetation
[edit]The density of vegetation in mangrove forests, seagrass meadows, and tidal marshes is an important factor in carbon burial rates.[47][48] The density of the vegetation must be sufficient to change water flows enough to reduce erosion and increase sediment deposition.[49][47]</ref>
Nutrient load
[edit]Increases in carbon capture and sequestration have been observed in both mangrove and seagrass ecosystems which have been subjected to high nutrient loads, either intentionally or due to waste from human activities.[50] Intentional fertilization has been used in seagrass meadow restoration. Perches for Seabirds are installed in the meadow and the bird droppings are the fertilizer source. The fetrilization allows fast growing varieties of seagrasses to establish and grow. The species composition of these meadows is markedly different than the original seagrass meadow, although after the meadow has been reestablished and fertilization terminated, the meadows return to a species composition that more closely resembles an undisturbed meadow.[51] Research done on mangrove soils from the Red_Sea have shown that increases in nutrient loads to these soils do not increase carbon mineralization and subsequent CO2 release.[52] This neutral effect of fertilization was not found to be true in all mangrove forest types. Carbon capture rates also increased in these forests due to increased growth rates of the mangroves. In forests with increases in respiration there were also increases in mangrove growth of up to six times the normal rate.[53]
Engineered approaches to blue carbon
[edit]A US Department of Energy study from 2001 proposed to replicate a natural process of carbon sequestration in the ocean by combining water rich in CO2 gas with carbonate [CO3-] to produce a bicarbonate [HCO3-] slurry. Practically, the engineered process could involve hydrating the CO2 from power plant flue gas and running it through a porous bed of limestone to ‘fix’ the carbon in a saturated bicarbonate solution. This solution could then be deposited at sea to sink in the deep ocean. The cost of this process, from capture to ocean burial, was estimated to range between $90 to $180 per tonne of CO2 and was highly dependent on the distance required to transport limestone, seawater, and the resulting bicarbonate solution.
Expected benefits from bicarbonate production over direct CO2 gas injection would be a significantly lesser increase in ocean acidity and a longer timescale for burial before the captured carbon would be released back to the atmosphere.[54]
References
[edit]- ^ a b c d Nelleman, C. "Blue carbon: the role of healthy oceans in binding carbon" (PDF).
- ^ a b c d McLeod, E. "A blueprint for blue carbon: toward an improved understanding of the role of vegetated coastal habitats in sequestering CO2" (PDF).
- ^ a b c d e Duarte, CM (2011). "Assessing the capacity of seagrass meadows for carbon burial: current limitations and future strategies". Ocean Coastal Management.
- ^ Greiner, Jill (2013). "Seagrass restoration enhances "blue carbon" sequestration in coastal waters". PLOS One.
- ^ Gattuso, J. (2006). "Light availability in the coastal ocean: impact on the distribution of benthic photosynthetic organisms and their contribution to primary production". Biogeosciences.
- ^ a b Fourqurean, James W. (2012). "Seagrass ecosystems as a globally significant carbon stock". Nature Geoscience.
- ^ Waycott, M (2009). "Accelerating loss of seagrasses across the globe threatens coastal ecosystems". Proceedings of the National Academy of Sciences of the USA (PNAS).
- ^ Bouillon, Steven (2008). "Mangrove production and carbon sinks: a revision of global budget estimates". Global Biogeochemical Cycles.
- ^ a b c Spaulding, MD (2010). "World atlas of mangroves" (PDF).
- ^ a b Alongi, Daniel M (2012). "Carbon sequestration in mangrove forests" (PDF). Future Science.
- ^ a b c d Alongi, DM (2002). "Present state and future of the world's mangrove forests" (PDF). Environmental Conservation.
- ^ Duarte, C.M. (2005). "Major rule of marine vegetation on the oceanic carbon cycle" (PDF). Biogeosciences.
- ^ a b c d e f g Chmura, Gail L (2003). "Global carbon sequestration in tidal, saline wetland soils" (PDF). Global Biogeochemical Cycles. Cite error: The named reference ":0" was defined multiple times with different content (see the help page).
- ^ Chmura, Gail L (2013). "What do we need to assess the sustainability of the tidal salt marsh carbon sink?". Ocean and Coastal Management.
- ^ Mudd, Simon, M. (2009). "Impact of dynamic feedbacks between sedimentation, sea-level rise, and biomass production on near-surface marsh stratigraphy and carbon accumulation". Estuarine, Coastal and Shelf Science.
{{cite journal}}: CS1 maint: multiple names: authors list (link) - ^ a b c d e f g Adam, Paul (2002). "Saltmarshes in a time of change". Environmental Conservation. Cite error: The named reference ":1" was defined multiple times with different content (see the help page).
- ^ a b Kumar, K., Dasgupta, C. N., Nayak, B., Lindblad, P., & Das, D. (2011). Development of suitable photobioreactors for CO 2 sequestration addressing global warming using green algae and cyanobacteria. Bioresource technology, 102(8), 4945-4953.
- ^ Kumar, K., Banerjee, D., & Das, D. (2014). Carbon dioxide sequestration from industrial flue gas by Chlorella sorokiniana. Bioresource technology,152, 225-233.
- ^ a b c Chung, I. K., Beardall, J., Mehta, S., Sahoo, D., & Stojkovic, S. (2011). Using marine macroalgae for carbon sequestration: a critical appraisal.Journal of Applied Phycology, 23(5), 877-886.
- ^ a b Bird, M. I., Wurster, C. M., de Paula Silva, P. H., Bass, A. M., & De Nys, R. (2011). Algal biochar–production and properties. Bioresource technology,102(2), 1886-1891.
- ^ Mcleod, E., Chmura, G. L., Bouillon, S., Salm, R., Björk, M., Duarte, C. M., ... & Silliman, B. R. (2011). A blueprint for blue carbon: toward an improved understanding of the role of vegetated coastal habitats in sequestering CO2.Frontiers in Ecology and the Environment, 9(10), 552-560.
- ^ Chi, Z., O’Fallon, J. V., & Chen, S. (2011). Bicarbonate produced from carbon capture for algae culture. Trends in biotechnology, 29(11), 537-541.
- ^ Chi, Z., Xie, Y., Elloy, F., Zheng, Y., Hu, Y., & Chen, S. (2013). Bicarbonate-based integrated carbon capture and algae production system with alkalihalophilic cyanobacterium. Bioresource technology, 133, 513-521.
- ^ Chung, I. K., Oak, J. H., Lee, J. A., Shin, J. A., Kim, J. G., & Park, K. S. (2013). Installing kelp forests/seaweed beds for mitigation and adaptation against global warming: Korean Project Overview. ICES Journal of Marine Science: Journal du Conseil, fss206.
- ^ Cite error: The named reference
:53was invoked but never defined (see the help page). - ^ a b Zhang, J. P., Cheng-De, S. H. E. N., Hai, R., Jun, W. A. N. G., & Wei-Dong, H. A. N. (2012). Estimating change in sedimentary organic carbon content during mangrove restoration in southern China using carbon isotopic measurements. Pedosphere, 22(1), 58-66.
- ^ a b Greiner, J. T., McGlathery, K. J., Gunnell, J., & McKee, B. A. (2013). Seagrass restoration enhances “blue carbon” sequestration in coastal waters. PloS one, 8(8), e72469.
- ^ Cite error: The named reference
:52was invoked but never defined (see the help page). - ^ a b Zhang, J. P., Cheng-De, S. H. E. N., Hai, R., Jun, W. A. N. G., & Wei-Dong, H. A. N. (2012). Estimating change in sedimentary organic carbon content during mangrove restoration in southern China using carbon isotopic measurements. Pedosphere, 22(1), 58-66.
- ^ a b Greiner, J. T., McGlathery, K. J., Gunnell, J., & McKee, B. A. (2013). Seagrass restoration enhances “blue carbon” sequestration in coastal waters. PloS one, 8(8), e72469.
- ^ * Fourqurean, James W., Joseph C. Zieman, and George V. N. Powell. 1992. “Phosphorus Limitation of Primary Production in Florida Bay: Evidence from C:N:P Ratios of the Dominant Seagrass Thalassia Testudinum.” ''Limnology and Oceanography'' 37 (1): 162–71. doi:10.4319/lo.1992.37.1.0162.
- ^ Mcleod, E., Chmura, G. L., Bouillon, S., Salm, R., Björk, M., Duarte, C. M., ... & Silliman, B. R. (2011). A blueprint for blue carbon: toward an improved understanding of the role of vegetated coastal habitats in sequestering CO2.Frontiers in Ecology and the Environment, 9(10), 552-560.
- ^ Chi, Z., Xie, Y., Elloy, F., Zheng, Y., Hu, Y., & Chen, S. (2013). Bicarbonate-based integrated carbon capture and algae production system with alkalihalophilic cyanobacterium. Bioresource technology, 133, 513-521.
- ^ Pendleton, Linwood; Donato, Daniel C.; Murray, Brian C.; Crooks, Stephen; Jenkins, W. Aaron; Sifleet, Samantha; Craft, Christopher; Fourqurean, James W.; Kauffman, J. Boone. "Estimating Global "Blue Carbon" Emissions from Conversion and Degradation of Vegetated Coastal Ecosystems". PLoS ONE. 7 (9). doi:10.1371/journal.pone.0043542. PMC 3433453. PMID 22962585.
{{cite journal}}: CS1 maint: unflagged free DOI (link) - ^ Pendleton, Linwood; Donato, Daniel C.; Murray, Brian C.; Crooks, Stephen; Jenkins, W. Aaron; Sifleet, Samantha; Craft, Christopher; Fourqurean, James W.; Kauffman, J. Boone. "Estimating Global "Blue Carbon" Emissions from Conversion and Degradation of Vegetated Coastal Ecosystems". PLoS ONE. 7 (9). doi:10.1371/journal.pone.0043542. PMC 3433453. PMID 22962585.
{{cite journal}}: CS1 maint: unflagged free DOI (link) - ^ Pendleton, Linwood; Donato, Daniel C.; Murray, Brian C.; Crooks, Stephen; Jenkins, W. Aaron; Sifleet, Samantha; Craft, Christopher; Fourqurean, James W.; Kauffman, J. Boone. "Estimating Global "Blue Carbon" Emissions from Conversion and Degradation of Vegetated Coastal Ecosystems". PLoS ONE. 7 (9). doi:10.1371/journal.pone.0043542. PMC 3433453. PMID 22962585.
{{cite journal}}: CS1 maint: unflagged free DOI (link) - ^ Pendleton, Linwood; Donato, Daniel C.; Murray, Brian C.; Crooks, Stephen; Jenkins, W. Aaron; Sifleet, Samantha; Craft, Christopher; Fourqurean, James W.; Kauffman, J. Boone. "Estimating Global "Blue Carbon" Emissions from Conversion and Degradation of Vegetated Coastal Ecosystems". PLoS ONE. 7 (9). doi:10.1371/journal.pone.0043542. PMC 3433453. PMID 22962585.
{{cite journal}}: CS1 maint: unflagged free DOI (link) - ^ Orth, Robert J.; Carruthers, Tim J. B.; Dennison, William C.; Duarte, Carlos M.; Fourqurean, James W.; Heck, Kenneth L.; Hughes, A. Randall; Kendrick, Gary A.; Kenworthy, W. Judson (2006-12-01). "A Global Crisis for Seagrass Ecosystems". BioScience. 56 (12): 987–996. doi:10.1641/0006-3568(2006)56[987:AGCFSE]2.0.CO;2. ISSN 0006-3568.
- ^ http://wwf.panda.org/about_our_earth/blue_planet/coasts/mangroves/mangrove_threats/)
- ^ "IUCN - Mangrove forests in worldwide decline". www.iucn.org. Retrieved 2016-02-29.
- ^ Macreadie, Peter I.; Hughes, A. Randall; Kimbro, David L. "Loss of 'Blue Carbon' from Coastal Salt Marshes Following Habitat Disturbance". PLoS ONE. 8 (7). doi:10.1371/journal.pone.0069244. PMC 3704532. PMID 23861964.
{{cite journal}}: CS1 maint: unflagged free DOI (link) - ^ Macreadie, Peter I.; Hughes, A. Randall; Kimbro, David L. "Loss of 'Blue Carbon' from Coastal Salt Marshes Following Habitat Disturbance". PLoS ONE. 8 (7). doi:10.1371/journal.pone.0069244. PMC 3704532. PMID 23861964.
{{cite journal}}: CS1 maint: unflagged free DOI (link) - ^ Macreadie, Peter I.; Hughes, A. Randall; Kimbro, David L. "Loss of 'Blue Carbon' from Coastal Salt Marshes Following Habitat Disturbance". PLoS ONE. 8 (7). doi:10.1371/journal.pone.0069244. PMC 3704532. PMID 23861964.
{{cite journal}}: CS1 maint: unflagged free DOI (link) - ^ Nittrouer, C. A. (2007). Continental margin sedimentation: From sediment transport to sequence stratigraphy. Malden, MA: Blackwell Pub. for the International Association of Sedimentologists.
- ^ Dandekar, P. (2012). Where Rivers Run Free. Retrieved February 24, 2016, from https://www.internationalrivers.org/resources/where-rivers-run-free-1670
- ^ "World's large river deltas continue to degrade from human activity". News Center. Retrieved 2016-02-24.
- ^ a b Cite error: The named reference
:13was invoked but never defined (see the help page). - ^ Cite error: The named reference
:23was invoked but never defined (see the help page). - ^ Hendriks, I. E., Sintes, T., Bouma, T. J., & Duarte, C. M. (2008). Experimental assessment and modeling evaluation of the effects of the seagrass Posidonia oceanica on flow and particle trapping.
- ^ Kumar, K., Banerjee, D., & Das, D. (2014). Carbon dioxide sequestration from industrial flue gas by Chlorella sorokiniana. Bioresource technology,152, 225-233.
- ^ Herbert, D. A., & Fourqurean, J. W. (2008). Ecosystem structure and function still altered two decades after short-term fertilization of a seagrass meadow. Ecosystems, 11(5), 688-700.
- ^ Keuskamp, J. A., Schmitt, H., Laanbroek, H. J., Verhoeven, J. T., & Hefting, M. M. (2013). Nutrient amendment does not increase mineralisation of sequestered carbon during incubation of a nitrogen limited mangrove soil.Soil Biology and Biochemistry, 57, 822-829.
- ^ Mcleod, E., Chmura, G. L., Bouillon, S., Salm, R., Björk, M., Duarte, C. M., ... & Silliman, B. R. (2011). A blueprint for blue carbon: toward an improved understanding of the role of vegetated coastal habitats in sequestering CO2.Frontiers in Ecology and the Environment, 9(10), 552-560.
- ^ Rau, G., K. Caldeira, K.G. Knauss, B. Downs, and H. Sarv, 2001. Enhanced Carbonate Dissolution as a Means of Capturing and Sequestering Carbon Dioxide. First National Conference on Carbon Sequestration Washington D.C., May 14-17, 2001.