
Anglerfish
| Anglerfish Temporal range:
| |
|---|---|

| |
| Humpback anglerfish, Melanocetus johnsonii | |
| Scientific classification | |
| Domain: | Eukaryota |
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Class: | Actinopterygii |
| Clade: | Percomorpha |
| Order: | Lophiiformes Garman, 1899 |
| Type species | |
| Lophius piscatorius | |

(B) Cryptopsaras couesii, 34.5 mm SL
(C) Himantolophus appelii, 124 mm SL
(D) Diceratias trilobus, 86 mm SL
(E) Bufoceratias wedli, 96 mm SL
(F) Bufoceratias shaoi, 101 mm SL
(G) Melanocetus eustalus, 93 mm SL
(H) Lasiognathus amphirhamphus, 157 mm SL
(I) Thaumatichthys binghami, 83 mm SL
(J) Chaenophryne quasiramifera, 157 mm SL.
The anglerfish are fish of the teleost order Lophiiformes (/ˌlɒfiɪˈfɔːrmiːz/).[1] They are bony fish named for their characteristic mode of predation, in which a modified luminescent fin ray (the esca or illicium) acts as a lure for other fish. The luminescence comes from symbiotic bacteria, which are thought to be acquired from seawater,[2][3] that dwell in and around the sea.
Some anglerfish are notable for extreme sexual dimorphism and sexual symbiosis of the small male with the much larger female, seen in the suborder Ceratioidei, the deep sea anglerfish. In these species, males may be several orders of magnitude smaller than females.[4]
Anglerfish occur worldwide. Some are pelagic (dwelling away from the sea floor), while others are benthic (dwelling close to the sea floor). Some live in the deep sea (such as the Ceratiidae), while others live on the continental shelf, such as the frogfishes and the Lophiidae (monkfish or goosefish). Pelagic forms are most often laterally compressed, whereas the benthic forms are often extremely dorsoventrally compressed (depressed), often with large upward-pointing mouths.[citation needed]
Evolution
[edit]The earliest fossils of anglerfish are from the Eocene Monte Bolca formation of Italy, and these already show significant diversification into the modern families that make up the order.[5] Given this, and their close relationship to the Tetraodontiformes, which are known from Cretaceous fossils, they likely originated during the Cretaceous.[6][7]
A 2010 mitochondrial genome phylogenetic study suggested the anglerfishes diversified in a short period of the early to mid-Cretaceous, between 130 and 100 million years ago.[8] A more recent preprint reduces this time to the Late Cretaceous, between 92 to 61 million years ago.[7] Other studies indicate that anglerfish only originated shortly after the Cretaceous-Paleogene extinction event as part of a massive adaptive radiation of percomorphs, although this clashes with the extensive diversity already known from the group by the Eocene.[7][9] A 2024 study found that all anglerfish suborders most likely diverged from one another during the Late Cretaceous and Paleocene, but the multiple families of deep-sea anglerfishes (Ceratioidei), as well as their trademark sexual parasitism, originated during the Eocene in a rapid radiation following the Paleocene-Eocene thermal maximum.[10]
Classification
[edit]Anglerfishes are classified by the 5th edition of Fishes of the World as set out below into 5 suborders and 18 families.[11][12] The following taxa have been arranged to show their evolutionary relationships.[4][13]
- Suborder Lophioidei Regan, 1912
- Family Lophiidae Rafinesque, 1810 (Monkfishes and goosefishes)
- Suborder Antennarioidei Regan, 1912
- Family Antennariidae Jarocki 1822 (Frogfishes)
- Family Tetrabrachiidae Regan, 1912 (Tetrabrachid frogfishes)
- Family Lophichthyidae Boeseman, 1964 (Lophichthyid frogfishes)
- Family Brachionichthyidae Gill, 1863 (Handfishes or warty anglerfishes)
- Suborder Chaunacoidei Pietsch & Grobecker, 1987
- Family Chaunacidae Gill, 1863 (Sea toad)
- Suborder Ogcocephaloidei Pietsch, 1984
- Family Ogcocephalidae Gill, 1893 (Batfishes)
- Suborder Ceratioidei Regan, 1912
- Family Caulophrynidae Goode & Bean, 1896 (Fanfins)
- Family Neoceratiidae Regan, 1926 (Spiny seadevils)
- Family Melanocetidae Gill, 1878 (Black seadevils)
- Family Himantolophidae Gill, 1861 (Footballfishes)
- Family Diceratiidae Regan & Trewavas, 1932 (Double anglers)
- Family Oneirodidae Gill, 1878 (Dreamers)
- Family Thaumatichthyidae Smith & Radcliffe, 1912 (Wolftrap anglers)
- Family Centrophrynidae Bertelsen, 1951 (Prickly seadevils)
- Family Ceratiidae Gill, 1861 (Warty seadevils)
- Family Gigantactinidae Boulenger, 1904 (Whipnose anglers)
- Family Linophrynidae Regan, 1925 (Leftvents)
The relationships of the suborders within Lophiiformes as set out in Pietsch and Grobecker's 1987 Frogfishes of the world: systematics, zoogeography, and behavioral ecology is shown below.[8]
| Lophiiformes |
| |||||||||||||||||||||||||||

_(16059517767).jpg)
It has been found in phylogenetic studies that both the Lophiiformes and the Tetraodontiformes nest within the Acanthuriformes and are so classified as clades within that taxon.[14]
Anatomy
[edit]All anglerfish are carnivorous and are thus adapted for the capture of prey. Ranging in color from dark gray to dark brown, deep-sea species have large heads that bear enormous, crescent-shaped mouths full of long, fang-like teeth angled inward for efficient prey-grabbing. Their length can vary from 2–18 cm (1–7 in), with a few types getting as large as 100 cm (39 in),[15] but this variation is largely due to sexual dimorphism, with females being much larger than males.[16] Frogfish and other shallow-water anglerfish species are ambush predators, and often appear camouflaged as rocks, sponges or seaweed.[17]
Anglerfish have a flap, or the illicium, towards the distal end of their body on their first of two dorsal fins which extends to the snout and acts as a luring mechanism where prey will approach in a face-to-face manner.[18] The illicium is moved back and forth by five distinct pairs of muscles: namely the shorter erector and depressor muscles that dictate movement of the illicial bone, along with inclinator, protractor, and retractor muscles that aid motion of the pterygiophore.[19]
Specifically considering Cryptopsaras couesii, this deep sea ceratioid anglerfish has unique rotational biomechanics in its musculature. The robust retractor and protractor muscles move in a winding pattern in opposite directions along the length of the pterygiophore, which exists in a deep longitudinal ridge along the skull.[19] Further, the long and thin inclinator of the deep sea ceratioid anglerfish allows for a distinctly wide range of anterior and posterior motion, assisting in the movement of the luring apparatus to aid in the ambush of prey.[19]
Most adult female ceratioid anglerfish have a luminescent organ called the esca at the tip of a modified dorsal ray (the illicium or fishing rod; derived from Latin ēsca, "bait"). The organ has been hypothesized to serve the purpose of luring prey in dark, deep-sea environments, but also serves to call males' attention to the females to facilitate mating.
The source of luminescence is symbiotic bacteria that dwell in and around the esca, enclosed in a cup-shaped reflector containing crystals, probably consisting of guanine. Anglerfish make use of these symbiotic relationships with extracellular luminous bacteria.[20] Atypical of luminous symbionts that live outside of the host's cells, the bacteria found in the lures of anglerfish are experiencing an evolutionary shift to smaller and less developed genomes (genomic reduction) assisted by transposon expansions.[20] Only a handful of luminescent symbiont species can associate with deep-sea anglerfishes.[3] In some species, the bacteria recruited to the esca are incapable of luminescence independent of the anglerfish, suggesting they have developed a symbiotic relationship and the bacteria are unable to synthesize all of the chemicals necessary for luminescence on their own. They depend on the fish to make up the difference. While females found within most anglerfish families have bioluminescence, there are exceptions including the Caulophrynidae and Neoceratiidae families.[21]
The bacterial symbionts are not found at consistent levels throughout stages of anglerfish development or throughout the different depths of the ocean.[22] Sequencing of larval organisms of the Ceratioidei suborder show an absence of bacterial symbionts, while sequencing of adult anglerfish showed higher levels of bioluminescent bacterial symbionts.[22] This correlates to the mesopelagic region having the highest levels of symbiont relationships in the anglerfish samples, as this is where adult anglerfish reside for most of their lives after their larval stage.[22] Electron microscopy of these bacteria in some species reveals they are Gram-negative rods that lack capsules, spores, or flagella. They have double-layered cell walls and mesosomes. A pore connects the esca with the seawater, which enables the removal of dead bacteria and cellular waste, and allows the pH and tonicity of the culture medium to remain constant. This, as well as the constant temperature of the bathypelagic zone inhabited by these fish, is crucial for the long-term viability of bacterial cultures.[23]
The light gland is always open to the exterior, so it is possible that the fish acquires the bacteria from the seawater. However, it appears that each species uses its own particular species of bacteria, and these bacteria have never been found in seawater. Haygood (1993) theorized that esca discharge bacteria during spawning and the bacteria are thereby transferred to the eggs.[23]
Some evidence shows that some anglerfish acquired their bioluminescent symbionts from the local environment. Genetic materials of the symbiont bacteria is found near the anglerfish, indicating that the anglerfish and their associated bacteria are most likely not evolved together and the bacteria take difficult journeys to enter the host.[3] In a study on Ceratioid anglerfish in the Gulf of Mexico, researchers noticed that the confirmed host-associated bioluminescent microbes are not present in the larval specimens and throughout host development. The Ceratioids likely acquired their bioluminescent symbionts from the seawater.[2] Photobacterium phosphoreum and members from kishitanii clade constitute the major or sole bioluminescent symbiont of several families of deep-sea luminous fishes.[24]
It is known that genetic makeup of the symbiont bacteria has undergone changes since they became associated with their host.[3] Compared to their free-living relatives, deep-sea anglerfish symbiont genomes are reduced in size by 50%. Reductions in amino acid synthesis pathways and abilities to utilize diverse sugars are found. Nevertheless, genes involved in chemotaxis and motility that are thought to be useful only outside the host are retained in the genome. Symbiont genome contains very high numbers of pseudogenes and show massive expansions of transposable elements. The process of genome reduction is still ongoing in these symbionts lineages, and the gene loss may lead to host dependence.[25]
In most species, a wide mouth extends all around the anterior circumference of the head, and bands of inwardly inclined teeth line both jaws. The teeth can be depressed so as to offer no impediment to an object gliding towards the stomach, but prevent its escape from the mouth.[26] The anglerfish is able to distend both its jaw and its stomach, since its bones are thin and flexible, to enormous size, allowing it to swallow prey up to twice as large as its entire body.[27]
Behavior
[edit]Swimming and energy conservation
[edit]In 2005, near Monterey, California, at 1,474 metres depth, an ROV filmed a female ceratioid anglerfish of the genus Oneirodes for 24 minutes. When approached, the fish retreated rapidly, but in 74% of the video footage, it drifted passively, oriented at any angle. When advancing, it swam intermittently at a speed of 0.24 body lengths per second, beating its pectoral fins in-phase. The lethargic behavior of this ambush predator is suited to the energy-poor environment of the deep sea.[28]
Another in situ observation of three different whipnose anglerfish showed unusual inverted swimming-behavior. Fish were observed floating inverted completely motionless with the illicium hanging down stiffly in a slight arch in front of the fish. The illicium was hanging over small visible burrows. It was suggested this is an effort to entice prey and an example of low-energy opportunistic foraging and predation. When the ROV approached the fish, they exhibited burst swimming, still inverted.[29]
The jaw and stomach of the anglerfish can extend to allow it to consume prey up to twice its size. Because of the limited amount of food available in the anglerfish's environment, this adaptation allows the anglerfish to store food when there is an abundance.[30]
Predation
[edit]
The name "anglerfish" derives from the species' characteristic method of predation. Anglerfish typically have at least one long filament sprouting from the middle of their heads, termed the illicium. The illicium is the detached and modified first three spines of the anterior dorsal fin. In most anglerfish species, the longest filament is the first. This first spine protrudes above the fish's eyes and terminates in an irregular growth of flesh (the esca), and can move in all directions. Anglerfish can wiggle the esca to make it resemble a prey animal, which lures the anglerfish's prey close enough for the anglerfish to devour them whole.[31] Some deep-sea anglerfish of the bathypelagic zone also emit light from their esca to attract prey.[32]
Because anglerfish are opportunistic foragers, they show a range of preferred prey with fish at the extremes of the size spectrum, whilst showing increased selectivity for certain prey. One study examining the stomach contents of threadfin anglerfish off the Pacific coast of Central America found these fish primarily ate two categories of benthic prey: crustaceans and teleost fish. The most frequent prey were pandalid shrimp. 52% of the stomachs examined were empty, supporting the observations that anglerfish are low energy consumers.[33]
Reproduction
[edit].gif)

Some anglerfish, like those of the Ceratiidae, or sea devils, employ an unusual mating method.[34] Because individuals are locally rare, encounters are also very rare. Therefore, finding a mate is problematic. When scientists first started capturing ceratioid anglerfish, they noticed that all of the specimens were female. These individuals were a few centimetres in size and almost all of them had what appeared to be parasites attached to them. It turned out that these "parasites" were highly dimorphic male ceratioids. This indicates some taxa of anglerfish use a polyandrous mating system. In some species of anglerfish, fusion between male and female when reproducing is possible due to the lack of immune system keys that allow antibodies to mature and create receptors for T-cells.[35] It is assumed they have evolved new immune strategies which compensate for the loss of B and T lymphocyte functions found in an adaptive immune system.[36]
Certain ceratioids rely on parabiotic reproduction. Free-living males and unparasitized females in these species never have fully developed gonads. Thus, males never mature without attaching to a female, and die if they cannot find one.[4] At birth, male ceratioids are already equipped with extremely well-developed olfactory organs that detect scents in the water. Males of some species also develop large, highly specialized eyes that may aid in identifying mates in dark environments. The male ceratioids are significantly smaller than a female anglerfish, and may have trouble finding food in the deep sea. Furthermore, growth of the alimentary canals of some males becomes stunted, preventing them from feeding. Some taxa have jaws that are never suitable or effective for prey capture.[37] These features mean the male must quickly find a female anglerfish to prevent death. The sensitive olfactory organs help the male to detect the pheromones that signal the proximity of a female anglerfish.
The methods anglerfish use to locate mates vary. Some species have minute eyes that are unfit for identifying females, while others have underdeveloped nostrils, making them unlikely to effectively find females by scent.[4] When a male finds a female, he bites into her skin, and releases an enzyme that digests the skin of his mouth and her body, fusing the pair down to the blood-vessel level.[36] The male becomes dependent on the female host for survival by receiving nutrients via their shared circulatory system, and provides sperm to the female in return. After fusing, males increase in volume and become much larger relative to free-living males of the species. They live and remain reproductively functional as long as the female lives, and can take part in multiple spawnings.[4] This extreme sexual dimorphism ensures that when the female is ready to spawn, she has a mate immediately available.[36] Multiple males can be incorporated into a single individual female with up to eight males in some species, though some taxa appear to have a "one male per female" rule.[4]
Symbiosis is not the only method of reproduction in anglerfish. In fact, many families, including the Melanocetidae, Himantolophidae, Diceratiidae, and Gigantactinidae, show no evidence of male symbiosis.[37] Females in some of these species contain large, developed ovaries and free-living males have large testes, suggesting these sexually mature individuals may spawn during a temporary sexual attachment that does not involve fusion of tissue. Males in these species also have well-toothed jaws that are far more effective in hunting than those seen in symbiotic species.[37]
Sexual symbiosis may be an optional strategy in some species of anglerfishes.[4] In the Oneirodidae, females carrying symbiotic males have been reported in Leptacanthichthys and Bertella—and others that were not still developed fully functional gonads.[4] One theory suggests the males attach to females regardless of their own reproductive development if the female is not sexually mature, but when both male and female are mature, they spawn then separate.[4]
| External videos | |
|---|---|
One explanation for the evolution of sexual symbiosis is that the relatively low density of females in deep-sea environments leaves little opportunity for mate choice among anglerfish. Females remain large to accommodate fecundity, as is evidenced by their large ovaries and eggs. Males would be expected to shrink to reduce metabolic costs in resource-poor environments and would develop highly specialized female-finding abilities. If a male manages to find a female, then symbiotic attachment is ultimately more likely to improve life-time fitness relative to free living, particularly when the prospect of finding future mates is poor. An additional advantage to symbiosis is that the male's sperm can be used in multiple fertilizations, as he always remains available to the female for mating. Higher densities of male-female encounters might correlate with species that demonstrate facultative symbiosis or simply use a more traditional temporary contact mating.[38]
The spawn of the anglerfish of the genus Lophius consists of a thin sheet of transparent gelatinous material 25 cm (10 in) wide and greater than 10 m (33 ft) long.[39] Such an egg sheet is rare among fish. The eggs in this sheet are in a single layer, each in its own cavity. The spawn is free in the sea. The larvae are free-swimming and have the pelvic fins elongated into filaments.[26]
Threats
[edit]Northwest European Lophius species are heavily fished and are listed by the ICES as "outside safe biological limits".[40] Additionally, anglerfish are known to occasionally rise to the surface during El Niño, leaving large groups of dead anglerfish floating on the surface.[40]
In 2010, Greenpeace International added the American angler (Lophius americanus), the angler (Lophius piscatorius), and the black-bellied angler (Lophius budegassa) to its seafood red list—a list of fish commonly sold worldwide with a high likelihood of being sourced from unsustainable fisheries.Red List Fish[41][42]
Human consumption
[edit]One family, the Lophiidae, is of commercial interest with fisheries found in western Europe, eastern North America, Africa, and East Asia. In Europe and North America, the tail meat of fish of the genus Lophius, known as monkfish or goosefish (North America), is widely used in cooking, and is often compared to lobster tail in taste and texture.
In Africa, the countries of Namibia and the Republic of South Africa record the highest catches.[43] In Asia, especially Japan, monkfish liver, known as ankimo, is considered a delicacy.[44] Anglerfish is especially heavily consumed in South Korea, where it is featured as the main ingredient in dishes such as Agujjim.
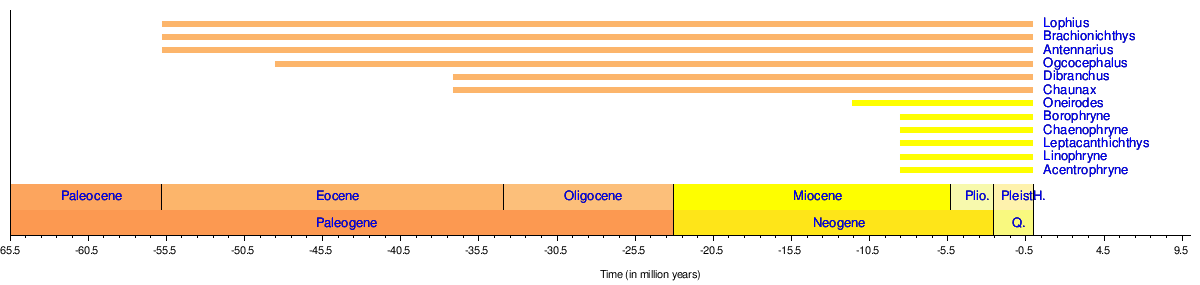
Timeline of genera
[edit]Anglerfish appear in the fossil record as follows:[45][46][47][48]
References
[edit]- ^ Froese, Rainer; Pauly, Daniel (eds.). "Order Lophiiformes". FishBase. February 2006 version.
- ^ a b Freed, Lindsay L.; Easson, Cole; Baker, Lydia J.; Fenolio, Danté; Sutton, Tracey T.; Khan, Yasmin; Blackwelder, Patricia; Hendry, Tory A.; Lopez, Jose V. (1 October 2019). "Characterization of the microbiome and bioluminescent symbionts across life stages of Ceratioid Anglerfishes of the Gulf of Mexico". FEMS Microbiology Ecology. 95 (10): fiz146. doi:10.1093/femsec/fiz146. ISSN 0168-6496. PMC 6778416. PMID 31504465.
- ^ a b c d Baker, Lydia J.; Freed, Lindsay L.; Easson, Cole G.; Lopez, Jose V.; Fenolio, Danté; Sutton, Tracey T.; Nyholm, Spencer V.; Hendry, Tory A. (1 October 2019). "Diverse deep-sea anglerfishes share a genetically reduced luminous symbiont that is acquired from the environment". eLife. 8: e47606. doi:10.7554/eLife.47606. ISSN 2050-084X. PMC 6773444. PMID 31571583.
- ^ a b c d e f g h i Pietsch, Theodore W. (25 August 2005). "Dimorphism, parasitism, and sex revisited: modes of reproduction among deep-sea ceratioid anglerfishes (Teleostei: Lophiiformes)". Ichthyological Research. 52 (3): 207–236. Bibcode:2005IchtR..52..207P. doi:10.1007/s10228-005-0286-2. S2CID 24768783.
- ^ Pietsch, Theodore W.; Carnevale, Giorgio (2011). "A New Genus and Species of Anglerfish (Teleostei: Lophiiformes: Lophiidae) from the Eocene of Monte Bolca, Italy". Copeia. 2011 (1): 64–71. doi:10.1643/CI-10-080. ISSN 0045-8511.
- ^ Chanet, Bruno; Guintard, Claude; Betti, Eric; Gallut, Cyril; Dettai, Agnes; Lecointre, Guillaume (1 September 2013). "Evidence for a close phylogenetic relationship between the teleost orders Tetraodontiformes and Lophiiformes based on an analysis of soft anatomy/Des donnees d'anatomie molle corroborent l'etroite parente entre Tetraodontiformes et Lophiiformes". Cybium. 37 (3): 179–199.
- ^ a b c Miller, Elizabeth Christina; Faucher, Rose; Hart, Pamela B.; Rincon-Sandoval, Melissa; Santaquiteria, Aintzane; White, William T.; Baldwin, Carole C.; Miya, Masaki; Betancur-R, Ricardo (30 October 2023), Phylogenomics reveals the deep ocean as an accelerator for evolutionary diversification in anglerfishes, doi:10.1101/2023.10.26.564281, retrieved 21 December 2023
- ^ a b Miya, M.; T. Pietsch; J. Orr; R. Arnold; T. Satoh; A. Shedlock; H. Ho; M. Shimazaki; M. Yabe (2010). "Evolutionary history of anglerfishes (Teleostei: Lophiiformes): a mitogenomic perspective". BMC Evolutionary Biology. 10 (1): 58. Bibcode:2010BMCEE..10...58M. doi:10.1186/1471-2148-10-58. PMC 2836326. PMID 20178642.
- ^ Alfaro, Michael E.; Faircloth, Brant C.; Harrington, Richard C.; Sorenson, Laurie; Friedman, Matt; Thacker, Christine E.; Oliveros, Carl H.; Černý, David; Near, Thomas J. (2018). "Explosive diversification of marine fishes at the Cretaceous–Palaeogene boundary". Nature Ecology & Evolution. 2 (4): 688–696. Bibcode:2018NatEE...2..688A. doi:10.1038/s41559-018-0494-6. ISSN 2397-334X. PMID 29531346.
- ^ Brownstein, Chase D.; Zapfe, Katerina L.; Lott, Spencer; Harrington, Richard; Ghezelayagh, Ava; Dornburg, Alex; Near, Thomas J. (2024). "Synergistic innovations enabled the radiation of anglerfishes in the deep open ocean". Current Biology. doi:10.1016/j.cub.2024.04.066. ISSN 0960-9822. PMID 38788708.
- ^ Nelson, J.S.; Grande, T.C.; Wilson, M.V.H. (2016). Fishes of the World (5th ed.). Hoboken, NJ: John Wiley & Sons. pp. 508–518. doi:10.1002/9781119174844. ISBN 978-1-118-34233-6. LCCN 2015037522. OCLC 951899884. OL 25909650M.
- ^ Richard van der Laan; William N. Eschmeyer & Ronald Fricke (2014). "Family-group names of recent fishes". Zootaxa. 3882 (2): 1–230. doi:10.11646/zootaxa.3882.1.1. PMID 25543675.
- ^ Arnold, Rachel J. (2014). Evolutionary Relationships of the Enigmatic Anglerfishes (Teleostei: Lophiiformes): Can Nuclear DNA Provide Resolution for Conflicting Morphological and Mitochondrial Phylogenies? (PhD thesis). University of Washington.
- ^ Near, Thomas J.; Thacker, Christine E. (2024). "Phylogenetic Classification of Living and Fossil Ray-Finned Fishes (Actinopterygii)". Bulletin of the Peabody Museum of Natural History. 65 (1): 3–302. doi:10.3374/014.065.0101.
- ^ "Anglerfish". National Geographic. 10 September 2010. Retrieved 28 February 2019.
- ^ "Fish Identification". fishbase.org. Retrieved 28 January 2019.
- ^ Pembury Smith, Matilda Q. R.; Ruxton, Graeme D. (2020). "Camouflage in predators". Biological Reviews. 95 (5): 1325–1340. doi:10.1111/brv.12612. hdl:10023/19948. ISSN 1464-7931. PMID 32410297. S2CID 218649415.
- ^ Yasugi, Masaki; Hori, Michio (June 2016). "Predominance of parallel- and cross-predation in anglerfish". Marine Ecology. 37 (3): 576–587. Bibcode:2016MarEc..37..576Y. doi:10.1111/maec.12309.
- ^ a b c Shimazaki, Mitsuomi; Nakaya, Kazuhiro (1 February 2004). "Functional anatomy of the luring apparatus of the deep-sea ceratioid anglerfish Cryptopsaras couesii (Lophiiformes: Ceratiidae)". Ichthyological Research. 51 (1): 33–37. Bibcode:2004IchtR..51...33S. doi:10.1007/s10228-003-0190-6. ISSN 1616-3915. S2CID 21508894.
- ^ a b Hendry, Tory A.; Freed, Lindsay L.; Fader, Dana; Fenolio, Danté; Sutton, Tracey T.; Lopez, Jose V. (5 July 2018). Moran, Nancy A. (ed.). "Ongoing Transposon-Mediated Genome Reduction in the Luminous Bacterial Symbionts of Deep-Sea Ceratioid Anglerfishes". mBio. 9 (3): e01033–18. doi:10.1128/mBio.01033-18. ISSN 2161-2129. PMC 6020299. PMID 29946051.
- ^ Ludt, William B.; Clardy, Todd R. (March 2022). "First detection of biofluorescence in a deep-sea anglerfish". Journal of Fish Biology. 100 (3): 843–846. Bibcode:2022JFBio.100..843L. doi:10.1111/jfb.14988. ISSN 0022-1112. PMID 34982469. S2CID 245670526.
- ^ a b c Freed, Lindsay L.; Easson, Cole; Baker, Lydia J.; Fenolio, Danté; Sutton, Tracey T.; Khan, Yasmin; Blackwelder, Patricia; Hendry, Tory A.; Lopez, Jose V. (2019). "Characterization of the microbiome and bioluminescent symbionts across life stages of Ceratioid Anglerfishes of the Gulf of Mexico". FEMS Microbiology Ecology. 95 (10). doi:10.1093/femsec/fiz146. PMC 6778416. PMID 31504465. Retrieved 26 March 2023.
- ^ a b Munk, Ole; Hansen, Kjeld; Herring, Peter J. (2009). "On the Development and Structure of the Escal Light Organ of Some Melanocetid Deep Sea Anglerfishes (Pisces: Ceratioidei)". Journal of the Marine Biological Association of the United Kingdom. 78 (4): 1321. doi:10.1017/S0025315400044520. ISSN 0025-3154. S2CID 84603596.
- ^ Haygood, Margo G.; Distel, Daniel L. (May 1993). "Bioluminescent symbionts of flashlight fishes and deep-sea anglerfishes form unique lineages related to the genus Vibrio". Nature. 363 (6425): 154–156. Bibcode:1993Natur.363..154H. doi:10.1038/363154a0. ISSN 0028-0836. PMID 7683390. S2CID 4346611.
- ^ Hendry, Tory A.; Freed, Lindsay L.; Fader, Dana; Fenolio, Danté; Sutton, Tracey T.; Lopez, Jose V. (26 June 2018). Moran, Nancy A. (ed.). "Ongoing Transposon-Mediated Genome Reduction in the Luminous Bacterial Symbionts of Deep-Sea Ceratioid Anglerfishes". mBio. 9 (3): e01033–18, /mbio/9/3/mBio.01033–18.atom. doi:10.1128/mBio.01033-18. ISSN 2150-7511. PMC 6020299. PMID 29946051.
- ^ a b One or more of the preceding sentences incorporates text from a publication now in the public domain: Chisholm, Hugh, ed. (1911). "Angler". Encyclopædia Britannica. Vol. 2 (11th ed.). Cambridge University Press. p. 15.
- ^ "Anglerfish". National Geographic. 10 September 2010. Retrieved 18 April 2018.
- ^ Luck, Daniel Garcia; Pietsch, Theodore W. (4 June 2008). "Observations of a Deep-sea Ceratioid Anglerfish of the Genus Oneirodes (Lophiiformes: Oneirodidae)". Copeia. 2008 (2): 446–451. doi:10.1643/CE-07-075. S2CID 55297852.
- ^ Moore, Jon A. (31 December 2001). "Upside-Down Swimming Behavior in a Whipnose Anglerfish (Teleostei: Ceratioidei: Gigantactinidae)". Copeia. 4. 2002 (4): 1144–1146. doi:10.1643/0045-8511(2002)002[1144:udsbia]2.0.co;2. JSTOR 1448539. S2CID 85724627.
- ^ "Deep Sea Anglerfish - Deep Sea Creatures on Sea and Sky".
- ^ Smith, William John (2009). The Behavior of Communicating: an ethological approach. Harvard University Press. p. 381. ISBN 978-0-674-04379-4.
Others rely on the technique adopted by a wolf in sheep's clothing—they mimic a harmless species. ... Other predators even mimic their prey's prey: anglerfish (Lophiiformes) and alligator snapping turtles Macroclemys temmincki can wriggle fleshy outgrowths of their fins or tongues and attract small predatory fish close to their mouths.
- ^ Piper, Ross (2007). Extraordinary Animals: An Encyclopedia of Curious and Unusual Animals, Greenwood Press.
- ^ Espinoza, Mario; Ingo Wehrtmann (2008). "Stomach content analyses of the threadfin anglerfish Lophiodes spilurus (Lophiiformes: Lophiidae) associated with deepwater shrimp fisheries from the central Pacific of Costa Rica". Revista de Biología Tropical. 4. 56 (4): 1959–70. doi:10.15517/rbt.v56i4.5772. PMID 19419094. Retrieved 4 October 2013.
- ^ Gorey, Colm (23 March 2018). "Scientists stunned to capture first mating footage of bizarre anglerfish". SiliconRepublic.com. Retrieved 23 March 2018.
- ^ Deep-sea anglerfishes have evolved a new type of immune system
- ^ a b c Isakov, Noah (2022). "Histocompatibility and Reproduction: Lessons from the Anglerfish". Life. 12 (1): 113. Bibcode:2022Life...12..113I. doi:10.3390/life12010113. ISSN 2075-1729. PMC 8780861. PMID 35054506.
- ^ a b c Pietsch, Theodore W. (8 March 1972). "A Review of the Monotypic Deep-Sea Anglerfish Family Centrophrynidae: Taxonomy, Distribution and Osteology". Copeia. 1972 (1): 17–47. doi:10.2307/1442779. JSTOR 1442779.
- ^ Miya, Masaki; Pietsch, Theodore W; Orr, James W; Arnold, Rachel J; Satoh, Takashi P; Shedlock, Andrew M; Ho, Hsuan-Ching; Shimazaki, Mitsuomi; Yabe, Mamoru; Nishida, Mutsumi (1 January 2010). "Evolutionary history of anglerfishes (Teleostei: Lophiiformes): a mitogenomic perspective". BMC Evolutionary Biology. 10 (1): 58. Bibcode:2010BMCEE..10...58M. doi:10.1186/1471-2148-10-58. PMC 2836326. PMID 20178642.
- ^ Prince, E. E. 1891. Notes on the development of the angler-fish (Lophius piscatorius). Ninth Annual Report of the Fishery Board for Scotland (1890), Part III: 343–348.
- ^ a b Clover, Charles (2004). The End of the Line: How overfishing is changing the world and what we eat. London: Ebury Press. ISBN 978-0-09-189780-2.
- ^ Greenpeace sea-food red list
- ^ Pietsch, Theodore W. (2009). Oceanic anglerfishes: extraordinary diversity in the deep sea. Berkeley: University of California Press. ISBN 978-0-520-94255-4. OCLC 1298208235.
- ^ Fariña, A. C; Azevedo, M; Landa, J; Duarte, R; Sampedro, P; Costas, G; Torres, M. A; Cañás, L (October 2008). "Lophius in the world: a synthesis on the common features and life strategies". ICES Journal of Marine Science. 65 (7): 1272–1280. doi:10.1093/icesjms/fsn140.
- ^ "The world's 50 best foods". CNN. CNN Travel. 12 July 2017. Retrieved 27 March 2023.
- ^ Sepkoski, Jack (2002). "A compendium of fossil marine animal genera". Bulletins of American Paleontology. 364: 560. Archived from the original on 23 July 2011. Retrieved 17 May 2011.
- ^ Carnevale, Giorgio; Theodore W. Pietsch; Gary T. Takeuchi; Richard W. Huddleston (2008). "Fossil Ceratioid Anglerfishes (Teleostei: Lophiformes) from the Miocene of the Los Angeles Basin, California" (PDF). Journal of Paleontology. 82 (5): 996–1008. Bibcode:2008JPal...82..996C. doi:10.1666/07-113.1. S2CID 129926776. Archived from the original (PDF) on 3 March 2016. Retrieved 27 October 2022.
- ^ Giorgio Carnevale; Theordore W. Pietsch (12 June 2009). "The deep-sea anglerfish genus Acentrophryne (Teleostei, Ceratioidei, Linophrynidae) in the Miocene of California" (PDF). Journal of Vertebrate Paleontology. 29 (2): 372–378. doi:10.1671/039.029.0232. ISSN 0272-4634. Wikidata Q114229338.
- ^ Nazarkin, Mikhail V.; Theodore W. Pietsch (2020). "A fossil dreamer of the genus Oneirodes (Lophiiformes: Ceratioidei) from the Miocene of Sakhalin Island, Russia". Geological Magazine. 157 (8): 1378–1382. Bibcode:2020GeoM..157.1378N. doi:10.1017/S0016756820000588. S2CID 225386060. Retrieved 27 October 2022.
Further reading
[edit]- Anderson, M. Eric, and Leslie, Robin W. 2001. Review of the deep-sea anglerfishes (Lophiiformes: Ceratioidei) of southern Africa. Ichthyological Bulletin of the J.L.B. Smith Institute of Ichthyology; No. 70. J.L.B. Smith Institute of Ichthyology, Rhodes University
External links
[edit]- Tree of Life web project: Lophiiformes
- Video (02:37) – Anglerfish mating on YouTube
- Lu, D. Anglerfish immune system lets them fuse with their mate. New Scientist 247, 19 (2020).
| International | |
|---|---|
| National | |